4.3.
Bacterial (microbial) lifestyle ...
...
oder das Leben kann so schön sein!
4.3.1.
Abiotische Wachstumsfaktoren
Bei
den Umgebungsfaktoren, die das bakterielle Leben und den bakteriellen Stoffwechsel
beeinflussen werden die Faktoren Temperatur, Wasserstoffionenkonzentration
(pH-Wert) des Milieus, osmotische Verhältnisse sowie Sauerstoff und
Kohlendioxyd Partialdruck in der gasförmigen Umgebung unterschieden. Einige
dieser Faktoren werden nachstehend beleuchtet.
4.3.1.1. Temperatur:
Man
unterscheidet in der Allgemeinen Mikrobiologie folgende Temperaturanpassungen
bei Bakterien:
à psychrophil, kältetolerante
Arten, die ein Temperaturoptimum bei
15 ºC aufweisen,
à mesophil, Bakterien mit einem
Temperaturoptimum bei 37 ºC (Abb.
13) und
à thermophil,
solche Bakterien, die an Extremstandorte angepasst
sind. Das Temperaturoptimum liegt
bei >55 ºC und kann im Extremfall 110 ºC erreichen.
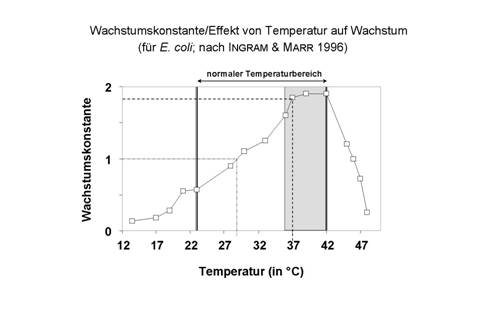
Abb. 13: Temperaturpräferenz von E. coli.
Die Temperaturpräferenz mesophiler
Bakterien stellt eine hervorragende Anpassung an die Körpertemperatur des
Menschen dar (Abb. 13). Medizinisch relevante Bakterienarten sind immer vom mesophilen Typus. Aber:
man kann mesophile Bakterien nichtsdestoweniger bei
tiefen und tiefsten Temperaturen (z.B. im flüssigem Stickstoff) einfrieren,
ohne das sie vollständig abgetötet werden. In der Regel sind auch lange Zeit
tief- oder tiefstgefrorene Bakterien wieder anzüchtbar
(dies trifft z.B. auf Erreger wie die Salmonellen zu, die an tiefgefrorenem Hühnerfleisch vorkommen und nach dem
Auftauen im Abtauwasser vermehrungsfähig sind)! Problematisch sind auch solche
Bakterien, die bei Kühlschranktemperaturen noch deutliche Vermehrung zeigen,
wie z.B. Listeria monocytogenes (Listerien-Embryopathie nach Infektion während der
Schwangerschaft möglich, Meningitis bei immunsupprimierten
Patienten).
4.3.1.2. Wasserstoffionenkonzentration (pH-Wert) des Milieus
Beispiele für Bakterien in neutraler und saurer
Umgebung. Der Großteil der medizinisch relevanten Bakterien bevorzugt die pH-neutrale Umgebung. Einige Bakterien sind jedoch auch auf
saures Milieu adaptiert bzw. stellen dieses Milieu durch ihre
Stoffwechselprodukte auf eine leicht sauren pH ein.
Beispiele sind die Lactobacillen in der Vaginalflora
(pH ca. 5) und Helicobacter pylori in der Magenschleimhaut (der
allerdings durch Stoffwechselprodukte eine Neutralisation seines Mikromilieus
erreicht).
4.3.1.3. Herkunft des
Zellkohlenstoffs und Energiestoffwechsel
Bakterien benötigen für ihren Gesamtstoffwechsel
Kohlenstoff. Dieser Kohlenstoff (in vielen Fällen Zucker) stellt den Anfangspunkt
des bakteriellen Energiestoffwechsels dar. Die Herkunft des Kohlenstoffs kann
sehr unterschiedlich sein, man unterscheidet autotrophe
und heterotrophe Ernährungstypen, je nach Herkunft
des Zellkohlenstoffs (Abb. 14):
Ø autotrophe (dabei
handelt es sich oft um phototrophe, d.h. Photosynthese
treibende Bakterien) Sie sind in der Lage, Zucker unter Ausnutzung der im
Sonnenlicht enthaltenen Energie direkt aus CO2 und H2O zu
assimilieren und die Lichtenergie so chemisch zu speichern.
Ø heterotrophe Bakterien benötigen eine externe Kohlenstoffquelle und
akquirieren den Zucker daher aus der Umgebung à die Zuckerverwertung (d.h. die Fähigkeit zur Verstoffwechselung ganz bestimmter Zucker à siehe lac-Operon) ist ein wesentliches Merkmal in der Differenzierung
von Bakterien im Routinelabor.

Abb. 14: Grundsätzliche Stoffwechseltypen von Bakterien.
4.4.
Stoffwechsel...
...
oder Katabolismus + Anabolismus
= Metabolismus
Der Gesamtstoffwechsel gliedert sich in die beiden Bereiche Katabolismus
(Energiegewinnung durch Oxidation von Kohlenstoffen à aerober Stoffwechsel; oder durch Gärung bzw. Fermentation von
Zucker à anaerober Stoffwechsel; Abb. 15) und den Anabolismus,
der Teil des Gesamtstoffwechsels, in dem aus einfachen Vorstufen biologische
Makromoleküle unter Energieverbrauch synthetisiert werden. Aerobe Bakterien
benötigen für die Oxidation des Zuckers Sauerstoff (und weisen eine höhere
Energieausbeute auf), während für anaerobe Bakterien der Sauerstoff sogar toxisch
sein kann. Anaerobier spalten den Zucker nur enzymatisch und weisen dabei eine sehr geringe
Energieausbeute auf. Bei dieser Gärung entstehen je nach Art der Spaltung
Gärungsendprodukte wie bspw. Ethanol. Bakterien, die den Menschen besiedeln
können sowohl vom anaeroben Stoffwechseltyp (Vorkommen z.B. in der Darmflora
oder am Grunde der Zahntaschen, als Infektionserreger auch in entzündlichen
Prozessen) wie auch vom aeroben Typ sein, einige Bakterien weisen in Bezug auf
ihre Präferenz gegenüber atmosphärischen Gasen besondere Anpassungen auf. So
bevorzugen Neisserien (u.a. Neisseria meningitidis; Meningokokken) oder Brucellen
(Erreger des undulierenden Fiebers) einen höhern Kohlendioxydanteil.
Aerobe
Bakterien verfügen über spezielle Enzyme, um im Stoffwechsel auftretende Sauerstoffmetaboliten abzufangen und unschädlich zu machen.
Diese Enzyme sind die Katalase sowie die Oxidase. Anaerobiern fehlen diese
Enzyme, so dass schädigende Sauerstoffmetaboliten
nicht abgefangen werden können.
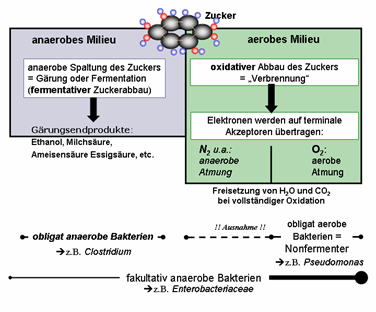
Abb. 15: Überblick über den katabolen
Stoffwechsel der Bakterien. Zucker als Energieträger und Kohlenstoffquelle kann
entweder oxydiert werden (höhere Energieausbeute: pro Mol Glucose etwa 38 Mol
ATP) oder anaerob gespalten werden (geringe Energieausbeute, 2-4 Mol ATP pro
Mol Glucose). Der aerobe Stoffwechsel setzt nicht notwendigerweise molekularen
Sauerstoff voraus. Wesentlich ist nur, dass Elektronen im Rahmen der Atmungskette
auf einen geeigneten Akzeptor übertragen werden. Auf
diese Weise können Bakterien auch in einer Sauerstofffreien Atmosphäre Zucker
oxydieren (anaerobe Atmung).
Bei der Wahl
des Zuckers, den Bakterien in ihren Stoffwechsel einschleusen können, ergeben
sich vielfältige Möglichkeiten und Anpassungen. Neben Bakterien, die überhaupt
keine Zucker als Kohlenstoffquelle nutzen gibt es solche, die nur einige wenige
Zucker verwerten können. Einige Bakterien können ihre enzymatische
Ausstattung zur Oxydation bestimmter Zucker in Abhängigkeit vom Angebot an
bestimmten Zuckern modulieren. Hierbei greifen elegante genetische Mechanismen.
So kann E. coli
z.B. mit Hilfe des sogenannten lac-Operons den
Stoffwechsel auf Laktose umstellen, wenn im umgebenden Milieu dieser Zucker in
einer bestimmten Konzentration verfügbar wird. Sinkt die Konzentration an
verfügbarer Laktose, können die Bakterien durch erneute genetische Umschaltung
wieder auf einen Stoffwechselweg für einen anderen Zucker umschalten.
Die
Überprüfung im Rahmen der Arbeiten des Routinelabors, ob bestimmte Stoffwechselleistungen
bei einem isolierten Erreger vorhanden sind oder nicht, stellt die Grundlage
der Differenzierung isolierter Krankheitserreger dar. So unterscheiden sich die
unterschiedlichen Erreger im Regelfall in dem komplexen Muster ihrer
Stoffwechselleistungen, zu denen sie befähigt sind. Wesentliche Unterschiede
ergeben sich z.B. in der Frage, welche Zucker verwertet werden können bzw. ob
bestimmte Enzyme der Atmungskette nachweisbar sind oder nicht (Katalase, Oxidase). So ist z.B.
der Erreger der eitrigen Gehirnhautentzündung, N. meningitidis, in der Lage in Kultur Maltose umzusetzen. Der
nahe verwandte und äußerlich kaum unterscheidbare Erreger des Trippers, Neisseria gonorrhoeae
kann nur Glucose umsetzten, so dass diese beiden Erreger durch die
Zuckerverwertung gut voneinander unterschieden werden können. Bei anderen
Bakterien kann mitunter die Abprüfung mehrerer stoffwechselphysiologischer
Leistungen notwendig werden. Hierfür gibt es vorgefertigte Miniaturtestsysteme
für den manuellen Einsatz (z.B. API Streifen[1])
als auch für den automatischen Einsatz (z.B. das Phoenix-System, welches nach
beimpfen eines „panels“, in dem
stoffwechselphysiologische Leistungen geprüft werden, eine automatische
Speziesbestimmung vornimmt). Gemeinsam mit den anderen Merkmalen wie Form der
Bakterienzelle und Anfärbbarkeit nach Gram oder
mittels spezieller Färbungen reflektieren die Unterschiede im
Stoffwechselverhalten auch die systematische Zugehörigkeit der Bakterien.
4.5. Zellteilung der
Bakterien...
...
und daraus resultierende Wachstumskurve
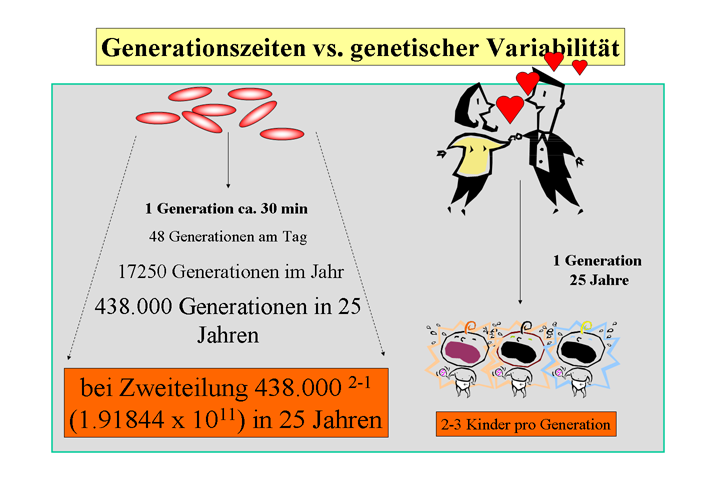
Die kurze Generationszeit der meisten medizinisch wichtigen Bakterien bringt den Vorteil der schnellen Anzüchtbarkeit im Labor und für die Bakterien den Vorteil, sich schnell an wechselnde Umweltbedingungen anpassen zu können. Die Generationszeiten der für die humanmedizinisch wichtigen Bakterien liegen in Kultur bei ca. 20 min (Escherichia coli; sichtbares
Wachstum auf festen Nährböden über Nacht) bis hin zu ca. 20 hrs
(Mycobacterium tuberculosis;
sichtbares Wachstum auf festen Nährböden nach 4 bis 8 Wochen). Schnelles
Wachstum ist die Grundlage für die Einteilung von Bakterien an Hand spezifischer
kultureller Merkmale und nur so kann eine schnelle Erregerdiagnostik erfolgen.
{kind=link}
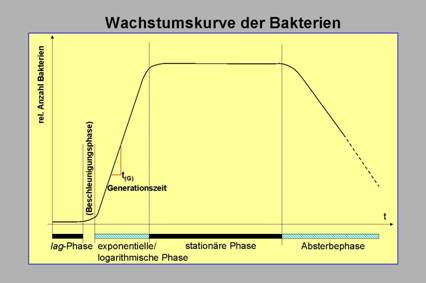
Abb. 16: Wachstumskurve der Bakterien. Ausgehend von der Annahme einer
frisch angesetzten Kultur zeigt die Kurve die Entwicklung der gesamten Population
in dieser Kultur.
Die Vorgänge
in einer Kultur lassen sich durch die Wachstumskurve für bakterielle Populationen beschreiben (Abb. 16). Man unterscheidet
die sogenannte (i)
lag-Phase, während derer die Anpassung der Bakterien
an das Milieu stattfindet, gefolgt von der (ii)
exponentiellen Wachstumsphase —gekennzeichnet durch schnelle Vermehrung unter
optimalen Wachstumsbedingungen (in dieser Phase wird die Generationszeit der
Bakterien bestimmt), die (iii)
stationäre Phase, in der keine makroskopische Zunahme
der Population zu beobachten ist, absterbende und durch Teilung neu hinzu
kommende Bakterien halten sich die Waage [makroskopischer
Stillstand, es setzen dichteabhängige Regulationsmechanismen ein] und
schlussendlich die (iv)
Absterbephase in der die Population auf Grund Überschreitens
der Nährstoffkapazität des Milieus und durch Anhäufung hemmender oder toxischer
Substanzen zu Grunde geht.
Die Kenntnis
dieser Wachstumskurve hat durchaus praktischen Wert, sowohl für die
Routinediagnostik als auch für die experimentelle Forschung, da man davon
ausgehen muss, dass sich Bakterien nur innerhalb der exponentiellen Wachstumsphase
„typisch“ in Bezug auf ihre
charakterisierenden Eigenschaften verhalten.
Wachstumskurve
für Populationen im Allgemeinen
(global und bezogen auf einzelne definierte Lebensräume):
Jeder
Lebensraum kann eine bestimmte Populationsgröße am Leben erhalten. Wird die
Populationsgröße kritisch, so greifen Regulationsmechanismen. Dies führt in
einer bakteriellen Kultur z.B. über die Anhäufung toxischer Produkte zum
Eintritt in die Absterbephase. In der menschlichen
Population bestimmter Regionen können Naturkatastrophen zum
Regulationsmechanismus für überbevölkerte Landstriche werden. In anderen
Bereichen kennzeichnen Unterernährung auf Grund mangelnder Ressourcen eine dem
Lebensraum unangepasste Populationsgröße. Erste theoretische Abhandlungen über
die Beziehung zwischen Bevölkerungswachstum und Vorhandensein von Ressourcen
wurden bereits von Thomas R. Malthus
in seinem „Bevölkerungsgesetz“ 1798 verfasst. Die moderne Populationsbiologie
spricht von dichteabhängigen Wachstum sowie dichteabhängigen Faktoren.